
Prof. Dr. Klaus Harter
Universität Tübingen
ZMBP, Pflanzenphysiologie
Auf der Morgenstelle 32
D-72076 Tübingen / Germany
klaus.harter@zmbp.uni-tuebingen.de
Tel.: ++49-7071/29-72605
Secretaries
Elke Fischer
Tue-Fri: 13:00-18:00
++49-7071/29-78802
elke.fischer@zmbp.uni-tuebingen.de
++49-7071/29-3287
Nicole Doppstadt
Thu: 9:00-12:30 / 14:00-18:30
Fri: 14:00-18:00
++49-7071/29-73225
pp@zmbp.uni-tuebingen.de
++49-7071/29-3287
Not occupied on Mondays.
Research Interests (Overview)
1. General Introduction
Plants face the problem that they are confined to their environment and have to endure and adapt to a broad array of environmental conditions. Furthermore, plants have to convert the "diffuse" energy source "light" into biochemical energy and to enrich mineral nutrients during uptake from the soil. To optimize photosynthesis and nutrient supply and, thus, growth and development in a highly competitive environment, plants depend on a multitude of signal perception systems for these endogenous and exogenous signals. Phytohormones, which are also perceived by specific receptors, fulfil a pivotal role in coordinating the responses to environmental cues with those induced by developmental processes. This complex coordination challenge does not lead to stochastic or chaotic reactions but to distinct output responses resulting in well-defined physiological adaptations as well as appropriate growth and development. These responses to a variety of simultaneously incoming exogenous and endogenous stimuli presuppose a kind of signal perception, transduction and processing machinery requiring highly networked molecular integration mechanisms. Although several specific and more-or-less linear signalling pathways have been described for higher plants in recent time, hardly anything is known about the components, composition and molecular mechanisms underlying signal integration.
Our present research focuses on the specific features of plant signal perception, signal transduction and information integration with our main concentration on the processes which are unique for higher plants.
These include:
- (i) The role of plant two-component systems in the perception, transduction and integration of exogenous and endogenous signals.
- (ii) Target genes and the post-translational regulation of the Arabidopsis subclass C basic leucine-zipper (bZIP) transcription factors and the analysis of their function in plant growth and development.
- (iii) The function of calcineurin B-like calcium sensors (CBL) and CBL-dependent kinases (CIPK) in calcium signalling (this research is carried out in collaboration with Prof. Dr. J. Kudla, Universitaet Muenster; for details see www.uni-muenster.de/Biologie.Botanik/agkudla/kudla.html).
- (iv) The identification of regulatory gene networks using bioinformatic approaches.
2. Plant Two-Component Systems
Recent investigations suggest that during evolution higher plants adapted and further developed a unique molecular signal processing mechanism which appears to be perfectly suited to accomplish the signal integration function proposed above: The Two-Component System (TCS, Figure 1).
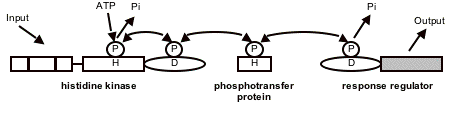
Figure 1.Basic features of the multistep two-component system (TCS) predominantly found in higher plants.
Signal (input) perception by the histidine kinase induces autophosphorylation of a conserved histidine (H) residue. The phosphoryl group (P) is relayed to a conserved Asp residue (D) that is localized in the C-terminal domain of the histidine kinase. The histidine-containing phosphotransfer proteins function as intermediates between the histidine kinase and the response regulator. The response regulator finally mediates the cellular output (adapted from Kakimoto 2003). For further details see Harter and Grefen (2004) in References.
In the model system Arabidopsis thaliana and other plant species, TCSs are mostly composed of hybrid histidine kinases, histidine phosphotransfer proteins and response regulators. It is noteworthy that the eukaryotic phytochrome photoreceptors have light-regulated two-component histidine kinases as ancestors.
The histidine kinases function as receptors for a specific endogenous or exogenous signal and - after autophosphorylation - transfer a phosphoryl residue in a strict one-to-one manner via a histidine phosphotransfer protein to a response regulator (Figure 1). Phosphorylation of response regulators induces their interaction with other proteins or binding to specific promoter regions of their target genes. In addition to this linear mode of action, the promiscuous interaction and histidine-to-aspartate phosphotransfer capability of TCS elements enable cross-talk between different signalling pathways, as we know from studies performed in prokaryotic systems, yeast and recently in plants. The histidine phosphotransfer proteins seem to play a crucial role, because of their ability to accept the phosphoryl groups from histidine kinases and to distribute them to response regulators.
In higher plants, canonical and non-canonical TCS elements have been shown or suggested to contribute to the regulation of several responses to endogenous and environmental stimuli and to participate in many signal transduction pathways. These include the perception and transduction of phytohormones (cytokinin, ethylene) and osmotic pressure as well as the regulation of phyllotaxis and circadian clock, the promotion of flowering and female gametophyte development. Furthermore, elements of TCS are able to adjust the responsiveness of plants to phytohormones and light.
On account of the multitude of elements, their molecular features and their way of action we propose that in higher plants TCSs may not (always) be the prevalent mechanisms for signal transduction but rather establish a complex network that is predominantly responsible for maintaining fine-tuning and cross-talk between signalling pathways and, therefore, contributes to the generation of overlapping physiological and developmental responses.
The analysis of the spatial and temporal composition of this network, its function in the perception, transduction and integration of environmental and endogenous signals as well as its dynamic adaptation will be a major challenge of our future research.
3. Basic Leucine-Zipper Transcription Factors
Transcription factors play crucial roles in almost all biological processes and are classified by their DNA-binding domain. The basic region/leucine zipper (bZIP) transcription factors have a basic region that binds DNA and a leucine zipper for homo- and heterodimerization (Figure 2).
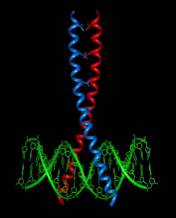
Figure 2. Schematic representation of a bZIP transcription factor binding to the major grove of double-stranded DNA (green). The basic domain contacts the DNA, whereas the leucine zipper mediates protein dimerization (source: www.icm.edu.pl).
Studies of a few plant bZIP transcription factors revealed that they participate in the regulation of diverse biological processes. Despite this recent progress, there is still little knowledge about the molecular and biochemical mechanisms of the post-translational regulation of plant bZIP factors.
In the course of our research we address the mechanisms of posttranslational modification (phosphorylation) and post-translational regulation of four bZIP transcription factors from Arabidopsis thaliana [namely Arabidopsis thaliana Common Plant Regulatory Factor 2-1 (AtCPRF2-1), AtCPRF2-2, AtCPRF2-3, AtCPRF2-4]. These factors form a small but distinct subfamily (group C) within the 75 members of Arabidopsis bZIP proteins. Our expression analyses and the evaluation of microarray data revealed that there is only minor regulation of the group C bZIP factors at transcriptional level. However, our preliminary data indicate that these factors are differentially regulated by post-translational mechanisms including intracellular distribution, dimerization and phosphorylation. We are currently determining the functional role of these post-translational mechanisms on the regulation of the activity of these bZIP transcriptional regulators.
In addition, we are interested in the in vivo target genes of the group C bZIP factors. We have , therefore, initiated a combination of different approaches which are aimed at identifying such target genes. Supported by results derived from physiological analysis of bZIP factor mutants and bZIP factor overexpressors, we expect to obtain insight into the function of these bZIP factors in growth and development or responses to environmental and endogenous stimuli in Arabidopsis.
4. Bioinformatic Approaches to Identify the Molecular Basis of Regulatory Gene Networks
It has long been considered that the evolution of regulatory interaction is a major driving force for the generation of evolutionary novelties. As more and more genomic sequences and expression data become available through public databases, we are now starting to resolve general issues such as, for instance, the dynamics of regulatory sequence evolution and the evolution of the specificity of transcriptional regulators.
We are interested in the comprehensive characterisation of cis-regulatory elements in the non-coding regions of genes. These cis-regulatory elements are recognized by transcription factors which actually trigger the activity of target genes. The more we understand how the position, orientation and combination of different cis-regulatory elements affect transcription, the more we will be able to predict expression patterns on the basis of promoter analysis.
For the broad range determination of gene expression profiles we are using microarray data from Arabidopsis thaliana. As a contributing laboratory of the multinational AtGenExpress project which is coordinated by the DFG-funded Arabidopsis Functional Genomic Network (AFGN), we have access to a multitude of expression profiles derived from various developmental stages and treatments of Arabidopsis plants (see Figure 3 as an example). Combining our present knowledge of physiological processes and signal transduction pathways, we aim to integrate these expression data into our current models of genome-wide gene expression.
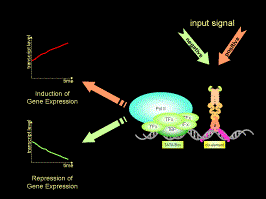
Figure 3. Regulatory principles and components involved in eukaryotic gene expression. For further details see text.
The availability of the complete genome sequences and the accumulating microarray data sets of the two model plant species Arabidopsis thaliana and Oryza sativa will also be a powerful tool for comparative analysis of the expression of orthologous gene pairs and might provide insights into cis-regulatory elements conserved for over 200 million years.
The regulation of gene expression is not restricted to the binding of transcription factors to cis-elements but also depends on the structural properties of the DNA neighbourhood. Thus, another focus of our bioinformatics approaches deals with the analysis of chromatin remodelling and epigenetics. Analysis of the factors and complexes involved in this type of gene expression control are not viable in most metazoan model organisms. Plants, therefore, may provide detailed information about chromatin-mediated processes impossible to attain in other eukaryotic system.
5. Methodical and Technical Aspects of our Research
At present our research is predominantly based on the model plant Arabidopsis thaliana but will be extended to other plant species (e.g. tobacco, tomato, rice). The majority of our methods are non-invasive or minimally-invasive. They include, for instance, in planta protein localisation using fluorescent protein markers, in planta protein-protein interaction (e.g. bimolekulare fluorescence complementation: BiFC; Figure 4) and in planta biochemical studies (e.g. protein phosphorylation) as well as the characterization of mutants created by both forward and reverse genetics. Comprehensive physiological analyses are accompanied by transcriptome and proteome studies. The non-invasive/minimally-invasive cell biological techniques are supported by in vitro biochemical methods (e.g. protein complex purification, CO-IP, in vitro BiFC, DNA/protein interaction) and in vivo approaches also performed in non-plant systems as for instance in yeast (one-, two-hybrid and split-ubiquitin assays , ternary protein complex formation). In summary, we use a wide array of state-of-the-art methods and techniques to perform our research on plant signal perception, signal transduction and signal integration.
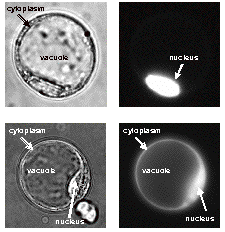
Figure 4. Dimerization of a nuclear (upper panel) and a cytoplasmic/nuclear (lower panel) proteins in living plant cells viewed by YFP-based bimolecular fluorescence complementation (BiFC). The interaction-proving YFP fluorescence images are presented at the right, whereas bright field images of the identical protoplasts are shown at the left. For further details see Walter et al. (2004) in References.
6. Acknowledgement
We thank the Deutsche Forschungsgemeinschaft for funding of our research.